Pterotrachea
Roger R. Seapy

- Pterotrachea coronata Niebuhr (in Forskål) 1775
- Pterotrachea hippocampus Phillipi 1836
- Pterotrachea scutata Gegenbaur 1855
Introduction
Species in the genus Pterotrachea have elongate and mostly cylindrical bodies, with the result that they are the most streamlined genus among the three heteropod families. The proboscis is slender, elongate and highly mobile, terminating in a small buccal mass (in contrast with the shorter and thicker proboscis in carinariids). The trunk is elongate and the cutis anterior to the swimming fin is thickened either ventro-laterally to form gelatinous folds (bib) or expanded laterally as an oval, cuticular disk. The tail is large and laterally compressed, which aids in burst swimming (resulting from side-to-side flexion of the trunk and tail). The tail terminates in two small, leaf-like lobes and, emerging from between the lobes, a filamentous extension, that may or may not be present. The larval shell is cast off following metamorphosis. A small sucker on the swimming fin is present only in males.
Brief Diagnosis:
A genus in the family Pterotracheidae with the following characteristics:
- Body streamlined; elongate and basically cylindrical in shape
- Proboscis slender, with a small buccal mass, and is highly mobile
- Cutis thickened ventro-laterally or laterally on the anterior part of the trunk, forming either a bib or an cuticular disk, respectively
- Tail large and laterally flattened, aiding in burst swimming involving side-to-side body flexion
- Swimming fin sucker present only in males
Characteristics
- Body morphology
- Body streamlined, with an elongate trunk and laterally-flattened tail that facilitate burst swimming using side-to-side body flexion (see title illustration)
- Proboscis slender, elongate and highly mobile, terminating in a small buccal mass (see title illustration)
- Trunk cutis anterior to the swimming fin thickened either ventro-laterally as a gelatinous fold or bib (P. hippocampus and P. coronata; first photograph below) or laterally as a flattened cuticular disk (P. scutata; second photograph below)
Click on an image to view larger version & data in a new window
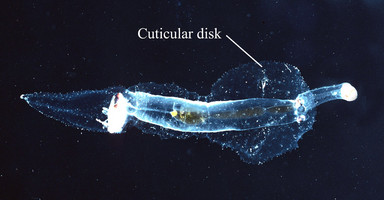
Figure. Left: Pterotrachea hippocampus viewed from the left side. Right: P. scutata in dorsal view. ©
- Viewed dorsally, eye shape either rectangular (P. coronata and P. scutata; first photograph below) or narrowly to broadly triangular, as a function of age (P. hippocampus; second and third photographs, respectively)
Click on an image to view larger version & data in a new window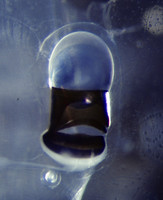


Figure. Eyes of Pterotrachea scutata (left) and P. hippocampus (center and right). ©
- Fin sucker small, located on ventral surface of swimming fin; present only in males. The sole function hypothesized for the male sucker has been to hold the female during mating. However, observations of a mating pair of P. hippocampus by Harbison (cited in Lalli and Gilmer, 1989) revealed that when the male and female were paired ventrally and head to head, the male's fin sucker contacted the female but was not seen to actually hold the female
Click on an image to view larger version & data in a new window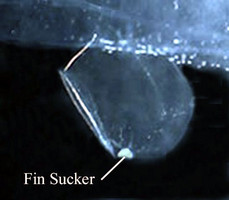
Figure. Swimming fin and fin sucker in male Pterotrachea coronata, viewed from the right side of the body. ©
- Tail terminates in two flattened lobes (see title illustration), between which an elongate filament emerges. The filament is comprised of pigmented nodules (see photograph below) and is present in both sexes. In the field R. Gilmer (cited in Lalli and Gilmer, 1989) observed that in P. coronata the filament can extend up to 60 mm in length and that it expands and contracts rapidly if the animal is disturbed. Unfortunately, during net capture the tail filament is often shed, and thus is missing in most preserved animals
Click on an image to view larger version & data in a new window
Figure. Tail filament of Pterotrachea coronata, viewed from right side. ©
- Roof of the buccal cavity bears two rows of additional palatine teeth; a feature unique to the genus and absent in all other heteropods (MacDonald, 1871)
- Shell present only in larvae (characterized below), as it is cast off following metamorphosis
- Tentacles absent in both males and females
- Radula with central rachidian teeth that are polycuspid, with a large, pointed median cusp and multiple side cusps (see photographs below). Lateral and marginal teeth are monocuspid
Click on an image to view larger version & data in a new window
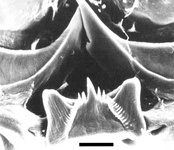
Figure. Scanning electron micrographs of central rachidian teeth in Pterotrachea hippocampus at low (left) and high (right) magnifications. Left photograph modified from Thiriot-Quiévreux (1973, Fig. 11) © 1973 C. Thiriot. Right photograph modified from Richter and Seapy (1999, Fig. 4D) © G. Richter
- Larvae of three types are recognized in the genus Pterotrachea, which were designated as larvae 1, 2 and 3 by Richter (1968)
- The shells of the three larvae all consist of about 2-1/2 whorls, but are distinguished by the following features (see photographs below): in larva 1, the surface is smooth and the last whorl separates from the inner whorls; in larva 2, the last whorl does not separate from the preceding whorls, the shell is thin and transparent, and the surface is tranversed by about 30 ridges; and, in larva 3 the shell is coiled and has a shape similar to larva 2, but the surface is smooth
Click on an image to view larger version & data in a new window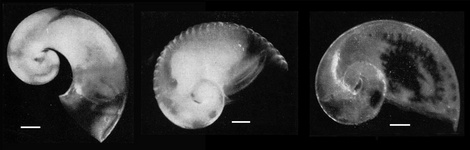
Figure. Shells (with retracted bodies) of the three larval types in the genus Pterotrachea: larvae 1 (left), 2 (center), and 3 (right). Plate created from photographs in Richter (1968, Figs. 19, 20, and 21). Scale bars = 100 µm. © 1968 G. Richter
- In her review of the Heteropoda, Thiriot-Quiévreux (1973) confirmed the identifications by Richter of the three larval types in the Mediterranean Sea (see photographs of larvae 1 and 2 below). She also reported that larva 2 was most frequently encountered
Similar to the larvae of Firoloida demarestia, larvae 1 and 2 possess a velum consisting of four long and slender lobes of equal length (see first photograph below). However, while three of the velar lobes in larva 3 are long and slender like those in the other two larvae, the left frontal lobe is widened and winglike (see second photograph below) and has a brown stain in the center of the tip (Richter, 1968)Click on an image to view larger version & data in a new window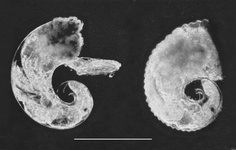
Figure. Photographs of Pterotrachea larvae 1 (left) and 2 (right). Scale bar = 500 µm. Modified from Thiriot-Quiévreux (1973, Fig. 7A,B). © 1973 C. Thiriot
Click on an image to view larger version & data in a new window
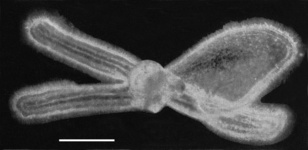
Figure. Photographs of the type 2 (left) and 3 (right) larvae of Pterotrachea. Modified from Richter (1968: Figs. 22,23). Scale bars = 50 µm © 1968 G. Richter
- Because the larval shell is shed following metamorphosis and early post-metamorphic development of the three species appears to be very similar, identification of the three larval types at the species level remains unresolved, although the first larva most probably belongs to P. hippocampus (see discussion in Comments below)
Comments
Krohn (1860) was the first to describe and illustrate the two larval types of Pterotrachea that Richter (1968) later termed Pterotrachea larvae 1 and 2.
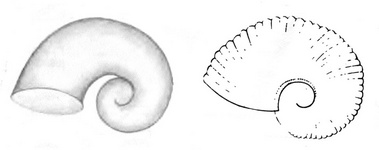
Figure. Two larval types of Pterotrachea, viewed from the left side. Modified from Krohn (1860, Figs. 22, 24). © 1860 A. Krohn
From western Mediterranean waters, Franc (1948) described a larva whose shell corresponded with that of Krohn's (1860) second larval type, and which he identified as Firola (Pterotrachea) coronata; although no justification was given for the species identification.
The two larval types of Krohn (1860) are also known from fossils, dated to the Quaternary Period (scanning electron micrographs below courtesy of Arie Janssen).
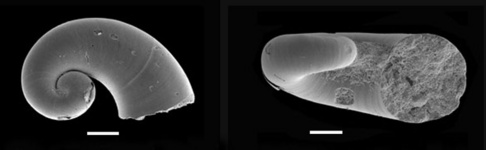

Figure. Scanning electron micrographs of fossil Pterotrachea larva 1 (above) and larva 2 (below), viewed from the right side (left) and aperture (right). Specimens from benthic core samples (700 m depth) in the Eastern Mediterranean and dated to the Quaternary Period; deposited in the National Natural History Museum Leiden, RGM 569.104a (top) and RGM 569.228a (bottom). Scale bars = 100 µm. © 2007 Arie W. Janssen
Richter (1968) made tentative species identifications of the three larval types based on observations of early development following metamorphosis. Only the early stages of eye formation provided a clue to species identity; the shape of the visceral nucleus, important in distinguishing P. coronata, was not discernable at this early stage of development. After metamorphosis of larva 1, the eye had a broad base, which Richter indicated would correspond with that seen in adult P. hippocampus. Following metamorphosis of larva 2, the base of the eye was found to be similar to the preceding species, but narrower; which Richter indicated was like that in adult P. minuta. Lastly, after metamorphosis in larva 3 the eye base was narrow, as seen in adult P. coronata or P. scutata. Given the determination that specimens previously identified as P. minuta are actually young P. hippocampus (Seapy, 1985, 2000), the tentative identity of larva 2 as P. minuta must be revisited. Richter's hypothesized identification of larva 1 as P. hippocampus is most probably correct, which leaves the other two larval types as either P. coronata or P. scutata. Unfortunately, the eyes of the latter two species are very similar in shape and appearance (see Seapy, 1985; Figs. 3F,G).
The three species in the genus Pterotrachea can be distinguished by the following:
| Species | Maximal body length | Cutis thckened on trunk anterior to swimming fin | Eye shape in dorsal view | Eye length to retinal width ratio (mean and range)* | Visceral nucleus shape | Visceral nucleus length to width ratio (mean and range)* |
|---|---|---|---|---|---|---|
| P. hippocampus | 80 mm | as ventro-lateral folds, forming a ventral bib | narrowly to broadly triangular | 1.3 (1.1-1.6) | short; tear-drop | 3.0 (2.0-3.7) |
| P. coronata | 330 mm | as ventro-lateral folds, forming a ventral bib | rectangular | 2.2 (1.9-2.5) | tall; narrow tear-drop | 5.6 (4.1-7.2) |
| P. scutata | 200 mm | as a laterally projecting, flattened oval disk | rectangular | 2.2 (2.1-2.4) | short; tear-drop | 3.0 (2.0-3.9) |
*Ratios (means and ranges) from Seapy (1985, Figs. 4 and 5). Based on regression analysis, the eye length to retinal width ratios of P. coronata and P. hippocampus decreased with increasing body length. A similar pattern was seen, but was less pronounced, for the ratio of visceral nucleus length to width in P. hippocampus. Remaining ratios showed essentially no change with increasing body length.
References
Franc, A. 1948. Véligères et mollusques gastéropodes des Baies d'Alger et de Banyuls. Journal de Conchyliologie (Paris) 88: 3-35.
Krohn, A. 1860. Beiträge zur Entwickelungsgeschichte der Pteropoden und Heteropoden. Verlag von Wilhelm Engelmann, Leipzig, pp. 1-43, 2 plates.
Lalli, C. M. and R. W. Gilmer. 1989. Pelagic snails. The biology of holoplanktonic gastropod mollusks. Stanford University Press, Stanford. 259 pp.
MacDonald, J. D. 1871. Examination of two soundings obtained in 62 and 68 fathoms respectively, latitude 41° 52', longitude 9° 8'; and the discovery of buccal teeth in the genus Firola. Quarterly Journal of Microscopic Science 11: 273-275.
Richter, G. 1968. Heteropoden und Heteropodenlarven im Oberflächenplankton des Golfs von Neapel. Pubblicazioni della Stazione Zoologica di Napoli 36: 346-400.
Richter, G. and R. R. Seapy. 1999. Heteropoda, pp. 621-647. In: D. Boltovskoy (ed.), South Atlantic Zooplankton. Backhuys Publishers, Leiden.
Seapy, R. R. 1985. The pelagic genus Pterotrachea (Gastropoda: Heteropoda) from Hawaiian waters: a taxonomic review. Malacologia 26(1-2): 125-135.
Seapy, R. R. 2000. Species discrimination among pelagic heteropods: resolution of the Pterotrachea hippocampus - P. minuta problem. Journal of Molluscan Studies 66: 99-117.
Thiriot-Quiévreux, C. 1973. Heteropoda. Oceanography and Marine Biology Annual Review 11: 237-261.
Title Illustrations

| Scientific Name | Pterotrachea coronata |
|---|---|
| Location | Hawaiian Islands |
| Specimen Condition | Live Specimen |
| Sex | Female |
| Life Cycle Stage | adult |
| View | left side |
| Image Use |
 This media file is licensed under the Creative Commons Attribution-NonCommercial License - Version 3.0. This media file is licensed under the Creative Commons Attribution-NonCommercial License - Version 3.0.
|
| Copyright |
©

|
About This Page
California State University, Fullerton, California, USA
Correspondence regarding this page should be directed to Roger R. Seapy at
Page copyright © 2007
 Page: Tree of Life
Pterotrachea .
Authored by
Roger R. Seapy.
The TEXT of this page is licensed under the
Creative Commons Attribution License - Version 3.0. Note that images and other media
featured on this page are each governed by their own license, and they may or may not be available
for reuse. Click on an image or a media link to access the media data window, which provides the
relevant licensing information. For the general terms and conditions of ToL material reuse and
redistribution, please see the Tree of Life Copyright
Policies.
Page: Tree of Life
Pterotrachea .
Authored by
Roger R. Seapy.
The TEXT of this page is licensed under the
Creative Commons Attribution License - Version 3.0. Note that images and other media
featured on this page are each governed by their own license, and they may or may not be available
for reuse. Click on an image or a media link to access the media data window, which provides the
relevant licensing information. For the general terms and conditions of ToL material reuse and
redistribution, please see the Tree of Life Copyright
Policies.
- First online 02 November 2007
- Content changed 19 September 2008
Citing this page:
Seapy, Roger R. 2008. Pterotrachea . Version 19 September 2008 (under construction). http://tolweb.org/Pterotrachea/28737/2008.09.19 in The Tree of Life Web Project, http://tolweb.org/