




Genistoid clade
Martin F. Wojciechowski and Johanna Mahn





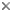
This tree diagram shows the relationships between several groups of organisms.
The root of the current tree connects the organisms featured in this tree to their containing group and the rest of the Tree of Life. The basal branching point in the tree represents the ancestor of the other groups in the tree. This ancestor diversified over time into several descendent subgroups, which are represented as internal nodes and terminal taxa to the right.

You can click on the root to travel down the Tree of Life all the way to the root of all Life, and you can click on the names of descendent subgroups to travel up the Tree of Life all the way to individual species.
For more information on ToL tree formatting, please see Interpreting the Tree or Classification. To learn more about phylogenetic trees, please visit our Phylogenetic Biology pages.
close boxIntroduction
The "Genistoid" clade is an informal name given to a group of papilionoids that includes the many genera traditionally classified in the tribes Genisteae, Brongniartieae, Crotalarieae, Euchresteae, Liparieae, Podalyrieae, and Thermopsideae, and some genera of the polyphyletic tribe Sophoreae (Käss and Wink, 1997; Crisp et al., 2000). The "genistoid alliance" as circumscribed by Polhill (1981, 1994) brought together this group of predominantly Old World, Southern Hemisphere tribes (and others) that have been considered closely related and comprising four separate lineages: one lineage included the predominantly Northern Hemisphere Genisteae sens. str., Euchresteae, Thermopsideae, together with the "Sophora" group of Sophoreae; the second consists of the mainly southern African Crotalarieae, Podalyrieae (now including Liparieae), and Hypocalypteae; a third comprised of the endemic Australasian Bossiaeeae and Mirbelieae; and the fourth, the Neotropical-Australian Brongniartieae.
Of the four lineages proposed by Polhill (1981, 1994), only the Australasian Bossiaeeae-Mirbelieae lineage appears to be monophyletic based on recent phylogenetic studies. Bossiaeeae plus Mirbelieae, together with the South African Hypocalypteae as its sister group, form a well-supported monophyletic group which is more closely related to Baphioid clade of Sophoreae (Kajita et al., 2001; Pennington et al., 2001) and the larger clade that includes the Millettioid sens. lat. clade and Hologalegina than to the Genistoids (Wojciechowski et al., 2004). The remaining three lineages (minus tribe Hypocalypteae) comprise the Genistoid clade; see Discussion of Phylogenetic Relationships below.
Characteristics
Members of the Genistoid clade are commonly shrubs and often have a broom-like habit, sharing a variety of morphological features, base chromosome numbers, and chemical synthetic capabilities (Polhill, 1994). The genistoids were previously distinguished by their different types and degrees of staminal fusion, but this characteristic has been shown to have multiple origins and proven unreliable as a defining characteristic (Crisp et al., 2000).
Common traits among species of genistoids include: xeromorphic growth, simple or reduced leaves or, if the leaves are compound, they have few leaflets, contracted vegetative structure and inflorescences, elaborated flowers, dimorphic anthers, and seeds that produce a hilar lobe extension of the radicle (Crisp et al., 2000). The synthesis of quinolizidine alkaloids may prove to be a chemical synapomorphy for this clade (see Discussion of Phylogenetic Relationships).
Discussion of Phylogenetic Relationships
Early molecular phylogenetic studies (e.g., Doyle et al., 1997; Käss and Wink, 1997) suggested the existence of a monophyletic group branching near the base of papilionoids that included many members of the Polhill's (1981) "genistoid alliance" together with certain genera of Sophoreae. Crisp et al. (2000) confirmed the polyphyly of the genistoid alliance and suggested the name be restricted to a smaller, well-supported "core genistoids" group that comprises the majority of the tribes from Africa and Eurasia (Crotalarieae, Euchresteae, Genisteae, Podalyrieae, Sophoreae sens. strict., and Thermopsideae). Studies based on molecular data alone or molecular data combined with phytochemical data (e.g., Crisp et al., 2000; Kajita et al., 2001; Pennington et al., 2001; Wink and Mohamed, 2003; Wojciechowski et al., 2004) are still far from complete with regard to resolving relationships among the members of these tribes within the larger groups that comprise this clade.
This clade has also received strong support from more recent studies which additionally suggest this core genistoids group is nested within a more inclusive Genistoid clade that includes the Brongniartieae (sensu Crisp and Weston, 1987), recently expanded to include two Neotropical genera, Cyclolobium Benth. and Poecilanthe Benth. (both formerly of tribe Millettieae), based on molecular sequence and phytochemical data (Crisp et al., 2000; Hu et al., 2000; Kite, 2005), and a number of largely woody, Neotropical genera of Sophoreae such as Acosmium Schott, Bolusanthus Harms, Diplotropis Benth., and most likely Ormosia Jackson and Clathrotropis Harms (Kajita et al., 2001; Pennington et al., 2001; Wojciechowski et al., 2004). The relationships among these particular taxa (e.g., Sophoreae pro parte) has not been definitively resolved by these studies but is the subject of active investigation.
The monophyly of the Genistoid clade is also supported by the taxonomic distribution of the synthesis of quinolizidine alkaloids (Kinghorn and Balandrin, 1984; Crisp et al., 2000; van Wyk, 2003); all species known to accumulate these particular alkaloids, with few exceptions (e.g. Calia, Kite and Pennington, 2003; and Ormosia, Kinghorn and Balandrin, 1984) are members of the Genistoid clade as defined here (Lewis et al., 2005). More comprehensive sampling for alkaloids and more robust molecular phylogenies of papilionoids should clarify the relationship of these specific taxa to the genistoids (i.e., as the sister group to genistoids) as well as resolve the issue of whether the synthesis of quinolizidine alkaloids is indeed a non-molecular synapomorphy for the Genistoid clade.
References
Crisp, M. D., and P. H. Weston. 1987. Cladistics and legume systematics, with an analysis of the Bossiaeeae, Brongniartieae, and Mirbelieae. Pages 65-130 in Advances in legume systematics, part 3 (C. H. Stirton, ed.). Royal Botanic Gardens, Kew, UK.
Crisp, M. D., S. Gilmore, and B.-E. Van Wyk. 2000. Molecular phylogeny of the genistoid tribes of papilionoid legumes. Pages 249-276 in Advances in legume systematics, part 9 (P. S. Herendeen and A. Bruneau, eds.). Royal Botanic Garden, Kew, UK.
Doyle, J. J., J. L. Doyle, J. A. Ballenger, E. E. Dickson, T. Kajita, and H. Ohashi. 1997. A phylogeny of the chloroplast gene rbcL in the Leguminosae: taxonomic correlations and insights into the evolution of nodulation. American J. Botany 84: 541-554.
Hu, J. M., M. Lavin, M. F. Wojciechowski, and M. J. Sanderson. 2000. Phylogenetic systematics of the tribe Millettieae (Leguminosae) based on matK sequences, and implications for evolutionary patterns in Papilionoideae. American J. Botany 87: 418-430.
Kajita, T., H. Ohashi, Y. Tateishi, C. D. Bailey, and J. J. Doyle. 2001. rbcL and legume phylogeny, with particular reference to Phaseoleae, Millettieae, and Allies. Systematic Botany 26: 515536.
Käss, E., and M. Wink. 1997. Phylogenetic relationships in the Papilionoideae (Family Leguminosae) based on nucleotide sequences of cpDNA (rbcL) and ncDNA (ITS1 and 2). Molecular Phylogenetics and Evolution 8:65-88.
Kinghorn, A. D., and M. F. Balandrin. 1984. Quinolizidine alkaloids in the Leguminosae: structural types, analysis, chemotaxonomy and biological activities. Pages 105-148 in Alkaloids: chemical and biological perspectives (S. W. Pelletier, ed.). John Wiley and Sons, New York, NY, USA.
Kite, G. C. 2005. Support for the removal of Cyclolobium from tribe Millettieae (Leguminosae) from its quinolizidine alkaloid status. Biochemical Systematics and Ecology 33: 39-43.
Kite, G. C., and R. T. Pennington. 2003. Quinolizidine alkaloid status of Styphnolobium and Cladrastis (Leguminosae). Biochemical Systematics and Ecology 31: 14091416.
Lewis, G., B. Schrire, B. MacKinder, and M. Lock (eds). 2005. Legumes of the world. Royal Botanical Gardens, Kew, UK.
Pennington, R. T., M. Lavin, H. Ireland, B. Klitgaard, J. Preston, and J.-M. Hu. 2001. Phylogenetic relationships of basal papilionoid legumes based upon sequences of the chloroplast trnL intron. Systematic Botany 26: 537556.
Polhill, R. M. 1981. Papilionoideae. Pages 191-208 in Advances in legume systematics, part 1 (R. M. Polhill and P. H. Raven, eds.). Royal Botanic Gardens, Kew, UK.
Polhill, R. M. 1994. Classification of the Leguminosae. Pages xxxv - xlviii in Phytochemical dictionary of the Leguminosae (F. A. Bisby, J. Buckingham, and J. B. Harborne, eds.). Chapman and Hall, New York, NY, USA.
Van Wyk, B.-E. 2003. The value of chemosystematics in clarifying relationships in the genistoid tribes of papilionoid legumes. Biochemical Systematics and Ecology 31: 875884.
Wink, M, and G. I. A. Mohamed. 2003. Evolution of chemical defense traits in the Leguminosae: mapping of distribution patterns of secondary metabolites on a molecular phylogeny inferred from nucleotide sequences of the rbcL gene. Biochemical Systematics and Ecology 31: 897-917.
Wojciechowski, M. F., M. Lavin, and M. J. Sanderson. 2004. A phylogeny of legumes (Leguminosae) based on analysis of the plastid matK gene resolves many well-supported subclades within the family. American J. Botany 91: 1846-1862.
Title Illustrations

| Scientific Name | Lupinus texensis Hook. |
|---|---|
| Copyright | © Bill Lindemann |

| Scientific Name | Sophora tomentosa L. |
|---|---|
| Copyright | © J. Paul Scurlock Collection |

| Scientific Name | Baptisia australis R.Br. |
|---|---|
| Specimen Condition | Live Specimen |
| Copyright | © 2006 |
About This Page
Martin F. Wojciechowski

Arizona State University, Tempe, Arizona, USA
Correspondence regarding this page should be directed to Martin F. Wojciechowski at and Johanna Mahn at
Page copyright © 2006 Martin F. Wojciechowski and
All Rights Reserved.
- First online 14 June 2006
- Content changed 14 June 2006
Citing this page:
Wojciechowski, Martin F. and Johanna Mahn. 2006. Genistoid clade. Version 14 June 2006. http://tolweb.org/Genistoid_clade/60334/2006.06.14 in The Tree of Life Web Project, http://tolweb.org/

{kind=link}