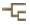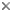Ommatidae
Frank Hünefeld and Rolf Georg Beutel



- Omma
- Tetraphalerus
- Brochocoleus

- Cionocoleus
- Eurydictyon
- Lithocupes
- Notocupes
- Notocupoides
- Rhabdocupes
- Zygadenia
Introduction
Ommatidae is a very small archostematan family with 2 extant genera. Omma comprises 4 species occuring in the eastern, south-eastern and south-western areas of Australia (O. mastersii, O. stanleyi, O. sagitta, and O. rutherfordi), and Tetraphalerus two described species (T. bruchi, T. wagneri) (Lawrence 1999; Hörnschemeyer 2005). In the Mesozoic, the family was moderately successful as documented by fossils from Jurassic and Cretaceous deposits in China, Siberia, Central Asia and western Europe (Tan et al. 2006). A considerable number of species were described and assigned to several extant and extinct genera (e.g., Tetraphalerus, †Amblomma, †Euryomma, †Odontomma, †Zygadenia). From their habitus, size and surface sculpture ommatid beetles are very similar to the closely related Cupedidae, but differ from them by the medially adjacent procoxae (prosternal process reduced) and abutting abdominal sternites. The species of Omma are known from densely forested coastal areas but also from drier Eucalyptus woodlands west of the great divide. The two species of Tetraphalerus live in in open, arid brushland (Hörnschemeyer 2005) in the southern half of South America.
Characteristics
Adults (Lawrence 1999; Hörnschemeyer 2005; Beutel et al. 2008; Friedrich et al. 2008)
The two gerena differ distinctly. The body is 6-27 mm long, more or less distinctly flattened (less so in Omma rutherfordi), elongate, and more or less parallel-sided (especially the Tetraphalerus species). The body surface is distinctly sculptured, with more or less specifically shaped tubercles, scale-like setae, and scales. The head is prognathous and moderately to strongly elongated. It is gradually or abruptly narrowed behind the compound eyes and a posterior collar region is present. Large dorsal protuberances or tubercles are present in O. rutherfordi and Tetraphalerus. The labrum is connate with the clypeus or fused with it. The antennae are inserted laterally in front of the moderately sized compound eyes. They are 11-segmented, filiform or moniliform, and at most as long as the head and thorax combined (reaching beyond the pronotal base in O. rutherfordi). The mandibles are characterised by 3 apical teeth arranged in a vertical row. A mola is lacking. The galea and lacinia are more or less equal, membranous and pubescent. The apical maxillary and labial palpomeres bear a sensory patch in a deep pit with a narrow (Omma) or wide opening. The prementum is large and plate like, covering the maxillary and labial lobes. The ligula is reduced. Internalised vestigial cervical sclerites are described for T. bruchi. The prothorax is slightly transverse to elongate (Tetraphalerus). Grooves for reception of the tarsi are absent. The well propleuron almost reaches the anterior margin of the segment and is usually not fused with the ventrite. The prosternal process is almost completely reduced and the procoxal cavities contiguous. The promesothoracic interlocking device is formed by a propleural cavity and a mesanepisternal process. A transverse ridge of the mesoventrite and a katepisternal mesocoxal joint are present. The mesofurcal arms arise with a common base. A distinct transverse ridge is present on the metaventrite and an exposed metatrochantin is present. The elytra bear 10 rows of cells or window punctures on the disc and one or more on the epipleura. The hind wings are well developed, with an oblongum cell and MP3 usually united with MP4. The posterior part is rolled in the resting position. The set of thoracic muscles is nearly complete and probably close to the coleopteran groundplan condition (Friedrich et al. 2008).
The abdomen is distinctly flattened dorsoventrally and approximately parallel-sided, with the exception of the last exposed ventrite (sternite VII). The dorsal side is completely covered by the elytra at rest. Eight tergites are present below the elytra in resting position. Tergites I – VI are semi-transparent and weakly sclerotized. Tergite VII is more strongly pigmented and sclerotized. Tergite VIII is strongly sclerotized and distinctly smaller than the preceeding ones; it is partially retracted below tergite VII in repose. Sternite I is greatly reduced and lacks any sclerotised elements. Sternite II, the first sclerotized ventrite, is completely covered by the metacoxae and firmly fused with sternite III. The terminal segments are retracted in repose. The terminalia are symmetrical and no torsion can be observed in repose or in copula. The anterolateral margins of tergite IX are produced into long sclerotized rods, the tergal apodemes, which are fused in the ventral midline. The aedeagus is trilobed and all elements of the copulatory apparatus are distinct. A distinctly developed, sclerotised basal piece is present. The parameres are deeply notched, with a larger, sclerotized proximal portion and a membranous distal part (autapomorphy of Ommatidae). The internal anatomy of the abdomen is only known for males of Tetraphalerus bruchi (Hünefeld et al., submitted). The muscular equipment is simple compared with other coleopteran lineages (18 muscles), in correlation with the symmetrical condition of the sclerites and the absence of a torsion of the genital apparatus. Almost all muscles occur pairwise and symmetrically. The testes are long, multi-coiled tubes like in other archostematans, Myxophaga (Torridincola) and Adephaga (F. Hünefeld, pers. obs.). The vasa deferentia are short and stout and connected to the unpaired ejaculatory duct directly in front of the aedeagus. A pair of accessory glands is present and shaped like elongated sacs. The testes, the vasa deferentia and the ejaculatory duct are enveloped by several layers of muscle fibres.
Larvae (Lawrence 1999; Beutel & Hörnschemeyer 2002a, b)
The immature stages of Ommatidae are largely unknown. One larval specimen collected east of Perth was described and tentatively assigned to the genus Omma. It is 12 mm long, elongated, subparallel, and gradually narrowing posteriorly. The body is slightly flattened. The cuticle is very lightly pigmented except for the mandibles, ligula, and anterolateral yellowish patches on the ventral prothoracic surface. The head is broad, slightly broader than long, and wedge-shaped in lateral view. A large, triangular emargination is present posterodorsally. The frontal and coronal sutures are not recognisable but a forked dorsal endocarina is present. Four well developed stemmata on both sides are equipped with individual cuticular lenses. The transverse labrum is movably attached to the clypeus. Its anterior margin bears a weak, rounded median lobe and a row of long setae. The epipharynx is covered with posteriorly directed hairs. A cluster of sensory pegs is present anteromesally and a transverse, slightly lunate sclerotisation at its base. The well developed tormae are strongly converging anteriorly and subparallel posteriorly. The antennae are moderately long, 4-segmented and anteriorly directed. They are inserted on a large antennifer. A long and slender sensorial appendage is inserted an antennomere 3. The mandibles are composed of a broad basal part and a strongly narrowed distal region with three apical teeth. A small retinaculum is present. The prominent mola lacks a raised margin. The maxillae are inserted in a deep fossa maxillaris. They are composed of a transverse, bipartite cardo, a short broad stipes, a 3-segmented palp inserted on a large palpifer, a palpomere-shaped galea with several stout spines, and a rounded lacinia. The fused submentum and mentum form a short, transverse plate with a pair of setae. The prementum is about as long as the maxillae and slightly narrowing anteriorly. It bears widely separated, small 2-segmented palps and a large and sclerotised ligula with a distinct anterior concavity. The hypostomal are convergent for the basal third and then divergent, extending almost to the posterior edge of the head capsule. A gula is not developed. The prothorax is less than half as long as wide. The pronotum is slightly sclerotised, rounded laterally and distinctly narrowing anteriorly. The prothoracic venter is not recognisably subdived. The meso- and metaterga are strongly transverse and equipped with transverse ampullae which lack asperities. The legs are 6-segmented (including claws), widely separated, and only about 0.15 x as long as the greatest mesothoracic width. The coxa is about half as long as wide and about as long as the trochanter, which is subequal to the tibia and tarsus and about 0.7 as long as the femur. The spiracles are vertically elongate. Abdominal segment I is slightly longer and about as wide as the metathorax. Tergites I and II are strongly transverse and slightly longer than the metatergite. Tergite III is also distinct but narrower and longer. The following tergites are less clearly defined. Segments IV-VIII are distinctly longer than I-III and narrower. VI and VII are almost parallel-sided. Segment IX is simple, posteriorly rounded, devoid of sclerotised processes, and indistinctly separated from X. Tergal ampullae and less strongly developed sternal ampullae are present on the first 2 or 3 segments. The anal opening is Y-shaped and surrounded by a dorsal and paired ventrolateral lobes. The abdominal spiracles are annular.
Discussion of Phylogenetic Relationships
Ommatidae was placed differently in several recently published phylogenetic studies with a focus on Archostemata. The adult mandibles with three teeth arranged in a vertical row and characters of the male genitalia suggest a clade comprising Ommatidae and the highly specialised Micromalthidae (Hörnschemeyer 2009). Other features support a sistergroup relationship between Ommatidae and the remaining extant and extinct groups of Archostemata (Beutel & Hörnschemeyer 2002a, b; Beutel & Friedrich 2008; Beutel et al. 2008). Several plesiomorphies are preserved in the presumptive larva of Omma (Lawrence 1999), with the corresponding apomorphic states in Cupedidae and Micromalthus (head slightly wider than long/head distinctly transverse, four stemmata/one eyespot or eyes absent, antenna moderately long/very short, margin of mola absent/present, eversible lobes of segment IX absent/present, sclerotised process of tergum IX absent/present) (see Beutel & Hörnschemeyer 2002a, b).
Omma is probably closer to the groundplan of the family. Within the genus, O. stanleyi is likely the sistergroup of the remaining three species (Hörnschemeyer 2009). Curved parameres with a shortened distal portion and a distinctly shortened penis are potential synapomorphies of O. rutherfordi and O. mastersi. The distinctly modified body-shape and coloration of O. rutherfordi suggests mimicry of mutillid wasps (Lawrence 1999). Tetraphalerus is characterised by the more elongate and flattened body, mushroom-shaped tubercles, conspicuous proturances on the strongly elongated head, and ventral antennal furrows. Eleven extinct genera are known from China alone: †Sinocupes, †Amblomma, †Euryomma, †Odontomma, †Brochocoleus, †Monticupes, †Cionocoleus, †Zygadenia, †Forticupes, †Fuscicupes, and †Picticupes. Whether all of them are phylogenetically justified needs to be tested.
References
Beutel, R.G. and T. Hörnschemeyer 2002. Larval morphology and phylogenetic position of Micromalthus debilis LeConte (Coleoptera: Micromalthidae). Systematic Entomology 27 (2): 169-190.
Beutel, R.G. and T. Hörnschemeyer 2002. Description of the larva of Rhipsideigma raffrayi (Coleoptera, Archostemata), with phylogenetic and functional implications. European Journal of Entomology 99: 53-66.
Beutel, R.G., S.-Q. Ge and T. Hörnschemeyer 2008. On the head morphology Tetraphalerus, the phylogeny of Archostemata and the basal branching events in Coleoptera. Cladistics 23: 1-29.
Beutel, R.G. and F. Friedrich 2008. The phylogeny of Archostemata (Coleoptera) and new approaches in insect morphology. Entomologia Generalis 31 (2): 141-154.
Friedrich, F., B. D. Farell and R. G. Beutel. 2009. The thoracic morphology of Archostemata and the relationships of the extant suborders of Coleoptera (Hexapoda). Cladistics 25: 1-37.
Hörnschemeyer, T. 2005. 5. Archostemata Kolbe, 1908. Pp. 29-42 in R. G. Beutel and R. A. B. Leschen (eds.), Handbook of Zoology, Handbook of Zoology, Vol. IV Arthropoda: Insecta. Part 38. Coleoptera, Vol. 1: Morphology and Systematics (Archostemata, Adephaga, Myxophaga, Polyphaga (partim). Walter De Gruyter, Berlin, New York.
Lawrence, J. F. 1999. The Australian Ommatidae (Coleoptera: Archostemata): new species, larva and discussion of relatioships. Invertebrate Taxonomy 13: 369-390.
Lawrence, J. F. and A. F. Newton, Jr. 1995. Families and subfamilies of Coleoptera (with selected genera, notes, references and data on family-group names). Pp. 779- 1006 in J. Pakaluk and S. A. Slipinski (eds.), Biology, Phylogeny, and Classification of Coleoptera: Papers Celebrating the 80th Birthday of Roy A. Crowson. Muzeum i Instytut Zoologii PAN, Warszawa.
Tan, J., D. Ren, C. Shih, S.-Q. Ge 2006. New fossil beetles of the family Ommatidae (Coleoptera: Archostemata) from the Jehol Biota of China. Acta Geologica Sinica 80:474-485.
Information on the Internet
- Classification of Fossil Beetles. A. G. Ponomarenko and A. G. Kirejtshuk, Zoological Institute of the Russian Academy of Sciences.
- New fossil beetles of the family Ommatidae (Coleoptera: Archostemata) from the Jehol Biota of China [pdf] by Tan et al (2006)
Title Illustrations

| Scientific Name | Omma stanleyi (Ommatidae) |
|---|---|
| Image Use |
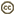 This media file is licensed under the Creative Commons Attribution License - Version 3.0. This media file is licensed under the Creative Commons Attribution License - Version 3.0.
|
| Copyright |
© 1996 David R. Maddison

|

| Scientific Name | Omma sagitta |
|---|---|
| Specimen Condition | Dead Specimen |
| Life Cycle Stage | Adult |
| View | Dorsal |
| Image Use |
This media file is licensed under the Creative Commons Attribution License - Version 3.0.
|
| Copyright |
© 1996 David R. Maddison
|

| Scientific Name | Tetraphalerus wagneri |
|---|---|
| Specimen Condition | Dead Specimen |
| Life Cycle Stage | Adult |
| View | Dorsal |
| Image Use |
This media file is licensed under the Creative Commons Attribution License - Version 3.0.
|
| Copyright |
© 1996 David R. Maddison
|
About This Page
Frank Hünefeld
Institut für Spezielle Zoologie und Evolutionsbiologie, Friedrich-Schiller-Universität
Rolf Georg Beutel
Friedrich-Schiller-Universität Jena, Germany
Correspondence regarding this page should be directed to Frank Hünefeld at and Rolf Georg Beutel at
Page copyright © 2011 Frank Hünefeld and
All Rights Reserved.
- Content changed 18 November 2010
Citing this page:
Hünefeld, Frank and Rolf Georg Beutel. 2010. Ommatidae. Version 18 November 2010 (under construction). http://tolweb.org/Ommatidae/8997/2010.11.18 in The Tree of Life Web Project, http://tolweb.org/